Article Text

Abstract
Background Cue reactivity, the enhanced sensitivity to conditioned cues, is associated with habitual and compulsive alcohol consumption. However, most previous studies in alcohol use disorder (AUD) compared brain activity between alcohol and neutral conditions, solely as cue-triggered neural reactivity.
Objective This study aims to find the neural subprocesses during the processing of visual alcohol cues in AUD individuals, and how these neural patterns are predictive for relapse.
Methods Using cue reactivity and rating tasks, we separately modelled the patterns decoding the processes of visual object recognition and reward appraisal of alcohol cues with representational similarity analysis, and compared the decoding involvements (ie, distance between neural responses and hypothesised decoding models) between AUD and healthy individuals. We further explored connectivity between the identified neural systems and the whole brain and predicted relapse within 6 months using decoding involvements of the neural patterns.
Findings AUD individuals, compared with healthy individuals, showed higher involvement of motor-related brain regions in decoding visual features, and their reward, habit and executive networks were more engaged in appraising reward values. Connectivity analyses showed the involved neural systems were widely connected with higher cognitive networks during alcohol cue processing in AUD individuals, and decoding involvements of frontal eye fields and dorsolateral prefrontal cortex could contribute to relapse prediction.
Conclusions These findings provide insight into how AUD individuals differently decode alcohol cues compared with healthy participants, from the componential processes of visual object recognition and reward appraisal.
Clinical implications The identified patterns are suggested as biomarkers and potential therapeutic targets in AUD.
Data availability statement
Data are available on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
What is already known on this topic
Alcohol cues triggered neural reactivity in alcohol use disorder (AUD) individuals.
What this study adds
This study disentangled the processing of alcohol cues into components of visual object recognition and reward appraisal.
How this study might affect research, practice or policy
The identification of the neural patterns related to componential processes may be helpful for precision medicine in AUD.
Background
Alcohol use disorder (AUD) is a major international public health issue with highly associated morbidity and mortality. It can be characterised as a disorder of neurocircuitry interacting with environmental and social factors.1
Cue reactivity is the enhanced sensitivity to conditioned cues. In AUD, these conditioned cues can trigger conditioned emotional or motivational reactions (ie, cue reactivity), which provide the basis for experiencing craving comprising the anticipation of reward or the occurrence of withdrawal symptoms in the case of not consuming the substance. In the literature, three models of cue reactivity have been proposed2: the conditioned withdrawal model, the conditioned compensatory response model and the conditioned appetitive-motivational model, which were recently unified in a framework of addiction3 4 where cue reactivity was conceptualised as the motivational change associated with addiction.
The neural activity triggered by cues was extensively reported and reviewed in previous publications. Existing neuroimaging evidence suggests that salient cues elicit increases in activity throughout the mesocorticolimbic system and nigrostriatal system.5 Activity in the mesocorticolimbic system, including the ventral tegmental area, ventral striatum, amygdala, anterior cingulate, prefrontal cortex, insula and hippocampus, as well as in sensory and motor cortices reflected the neural representations of reward values of cues and the motivational processes of incentive salience that guide drug-seeking behaviour. On the other hand, the nigrostriatal system is critical to habit learning and a transition from controlled to automatic behaviour, which consists primarily of dopamine projections from the substantia nigra to caudate and putamen (also referred to as the dorsal striatum) and globus pallidus. When increases in dorsal striatum cue reactivity were observed, the dorsal striatum circuits were also involved in the planning and execution of motor responses.5 Moreover, researchers found that sensory and motor functions could also importantly contribute to cue reactivity in addiction, and in both animals and humans, the activity of visual cortices could be modulated in viewing reward-mounted cues.6 In a study using an animal model of chronic alcohol drinking, a functional dedifferentiation in visual and sensorimotor networks was observed.7 Recently, a paper reviewed previous work in addition to research and demonstrated six large-scale brain networks (reward, habit, salience, executive, memory and self-directed networks) for understanding the dysfunctions in addictive disorders.8
Most of the previous studies compared brain activity between an alcohol condition and a neutral condition and reported cue-triggered brain activity as a contrast.5 9 However, comparing alcohol versus neutral cues usually only assesses the alcohol-cue-elicited activation, while the subprocesses leading to this change in the brain remain elusive. For example, how does the brain recognise the alcohol cues, and how are reward values represented in the brain? Until now, only a few studies considered the different reward values of alcohol cues in cue reactivity tasks, which could help to understand the process of reward appraisal (RA) in AUD individuals.
Objective
This study was designed to separately model processes of visual object recognition (VOR) and RA in cue reactivity and examine altered patterns of neural activity in AUD. Functional MRI (fMRI) data were examined with representational similarity analysis (RSA), a multivariate technique to model neural patterns.10 We hypothesised that, compared with healthy participants, AUD individuals show specific enhanced patterns in VOR and RA of the alcohol cues. Moreover, the level of enhancement of the neural patterns should be related to the clinical characteristics of AUD.
Methods
Participants
This study was based on a combined analysis of previous projects in AUD, which were designed with similar inclusion criteria and implemented with the same echo planar imaging sequence (for details, see online supplemental table 1). It comprised datasets from 238 (53 women) alcohol-dependent patients (Diagnostic and Statistical Manual of Mental Disorders, Fourth Edition and Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition criteria) and 229 (50 women) healthy participants recruited at the Central Institute of Mental Health, Mannheim, Germany, between 2008 and 2016. The average ages of AUD individuals and healthy participants were 47.0±10.8 and 45.7±12.7 years, respectively. Psychometric assessments included the Alcohol Dependence Scale, the Alcohol Urge Questionnaire (AUQ), the Obsessive Compulsive Drinking Scale (OCDS) and FORM 90 (for details, see online supplemental methods). The demographic and clinical overview of the participants are summarised in online supplemental table 2. All participants provided informed written consent according to the Declaration of Helsinki, and all projects in this study were approved by the local ethics committee of the University of Heidelberg (Ethics approval nos: 2007-095F-MA, 2009-215N-MA, 2010-348N-MA, 2011-303N-MA, 2009-215N-MA and 2015-540N-MA).
Supplemental material
Stimuli and experimental tasks
During the imaging session, visual alcohol and neutral stimuli were presented in blocks that were pseudo-randomised (tasks were described in online supplemental methods). The alcohol picture series was previously evaluated in a pilot study11 by AUD and healthy individuals with an attribute-rating task (outside the MRI scanner). In the task, the participants indicated their feeling towards these pictures from aspects of craving, valence and arousal (with three questions ‘when seeing this picture, how strong is your craving; how pleasant/unpleasant do you feel; how excited do you feel?’).
Imaging acquisition and preprocessing
Scanning was performed using a 3-T whole-body tomography scanner (MAGNETOM Trio with TIM technology; Siemens, Erlangen, Germany). T2*-weighted, echo planar images covering the entire brain were acquired. Imaging parameters were set to repetition time=2.41 s, echo time=25 ms, flip angle=80 degrees, number of slices=42, slice thickness=2 mm, voxel-gap=1 mm, voxel dimensions=3×3×3 mm3, field of view=192×192 mm2 and in-plane resolution=64×64. Visual stimuli were presented using Presentation software (Neurobehavioral Systems). fMRI data were processed and analysed using SPM12 (Wellcome Centre for Human Neuroimaging, University College London, UK). The first five scans were excluded from imaging analyses to avoid any artefacts caused by the effects of magnetic saturation. All images were realigned spatially, normalised to the SPM12 TPM MNI template, and unsmoothed images were used in the following analyses to preserve the fine spatial details in the fMRI signal.
Representational similarity analysis
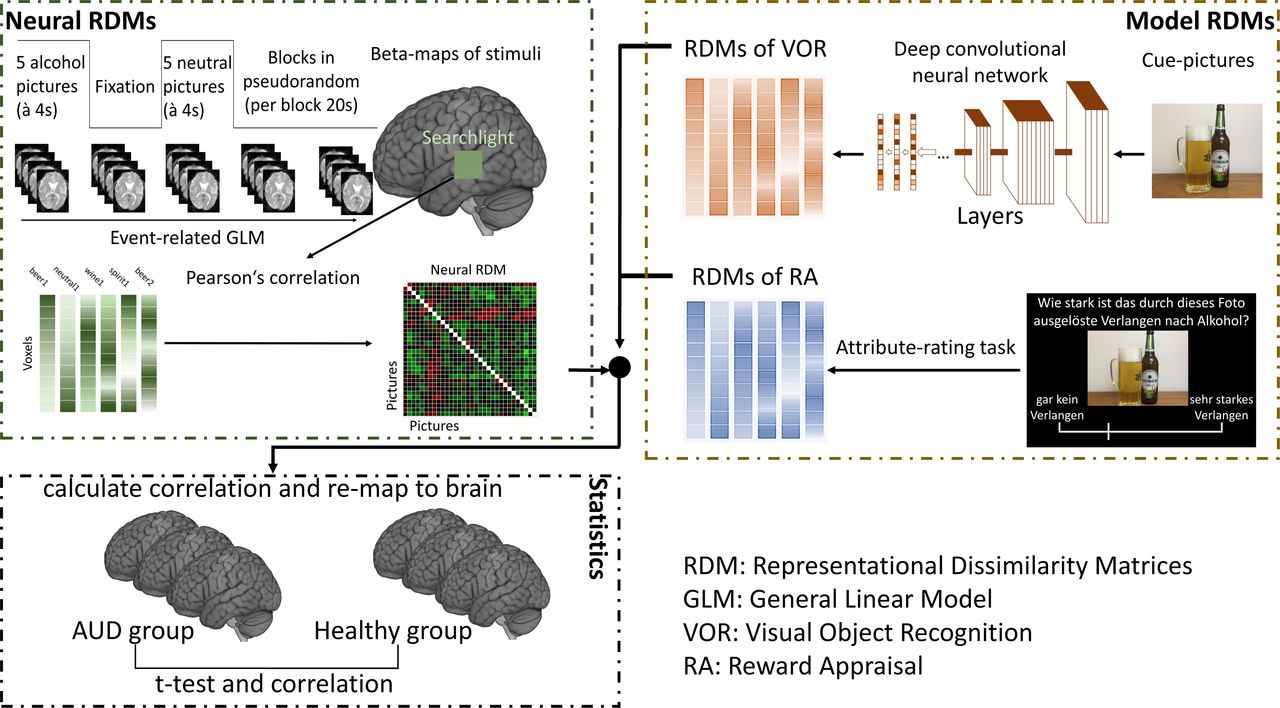
To find altered neural patterns in AUD individuals during the cue-reactivity task, we used RSA with two separate models of VOR and RA as shown in figure 1. Patterns of neural activity (neural representational dissimilarity matrices (RDMs)) were compared with model RDMs (VOR and RA) with a searchlight approach12 according to the hypotheses.

Procedure of representational similarity analysis with computational models. The functional MRI data were used to calculate the neural representational dissimilarity matrices (RDMs), while RDMS of visual object recognition (VOR) and reward appraisal (RA) were calculated with layers in deep convolutional neural network and ratings of pictures, respectively. Then patterns of neural activity (neural RDMs) were compared with model RDMs (VOR and RA) with a searchlight approach and followed with statistical tests. AUD, alcohol use disorder.
VOR and RA model
For the VOR, we used a pretrained deep convolutional neural network (dCNN) for model construction (for details, see online supplemental methods). We extracted the last three fully connected layers (fc8, fc7 and fc6) of the neural network for each picture and constructed the model RDMs of VOR. For the RDM of RA, we used ratings for each presented picture stimulus from a pilot study.11 The correlation distance (1−Pearson’s r) was used in distance calculation for model RDMs (figure 1).
Patterns of neural response
With preprocessed images (unsmoothed), we extracted voxel-wise fMRI responses (beta values) for all pictures using the general linear model in SPM12 including movement parameters. A previous study found block-designed fMRI tasks could also be modelled as event related and might even better explain neural responses.13 Therefore, though our cue-reactivity tasks in this study were block designed, event modelling was used in analyses to investigate the neural patterns with the hypothesised models. We then created individual RDMs based on the correlation distance for each pair of all the pictures.
Searchlight analysis
To quantify how well the different models were related to the neural patterns in the cue-reactivity task and assess the alerted involvement of brain regions in decoding information in AUD individuals, we correlated (Spearman’s rank correlation) each model RDM with individual neural RDM with a searchlight analysis (radius=10 mm). In this analysis, individual whole-brain maps of ‘decoding involvement’ (Fisher-transformed correlation coefficients) were obtained, which showed the similarity between the models and the brain response and were compared between AUD and healthy individuals.
Whole-brain connectivity to neural patterns of cue reactivity
To investigate the communication between the identified regions of neural representations of cue reactivity and other brain areas in AUD individuals, we performed psychophysiological interaction (PPI) analyses (for details, see online supplemental methods). The clusters with different neural representations identified by the group comparison of RSA were defined as seed regions (voxels in the clusters). The contrast ‘Alcohol versus Neutral’ was used in computing seed-to-voxel PPI in the whole brain.
Statistical analyses
The whole-brain decoding involvement maps of two groups were compared using SPM12 with a one-tailed two-sample t-test to find the enhanced neural pattern of AUD individuals. In the following analyses, a two-tailed one-sample t-test was used to examine the significant connectivity. To control for multiple comparisons, a voxel-wise threshold of p<0.0005 in combination with a cluster-extend threshold determined with random field theory in SPM12 was used for the cluster-corrected threshold of p<0.05. Based on the results of the between-group comparison, significant regions from both VOR and RA were defined as two neural patterns, and correlation analyses were conducted between the mean decoding involvements of the patterns and psychometric assessments in SPSS 25.
Relapse prediction with neural patterns
Finally, we conducted a prediction of relapse with the cue-reactivity neural patterns in a subsample of 59 AUD individuals, in whom follow-up data after cue-exposure–based extinction training and treatment-as-usual were available.14 The aim of prediction was relapse during a 6-month follow-up period. We extracted the mean ‘involvement value’ from all significant regions of interest (ROIs) (table 1) in the VOR and RA patterns separately as features and used a support vector machine (SVM) and lasso penalised logistic regression to predict the relapse (relapse=1 and abstinence=−1). The SVM model used a linear kernel and was trained with leave-one-out cross-validation. The weights of support vectors were used for investigating the most informative ROIs. The lasso penalised logistic regression model was trained with leave-one-out cross-validation, with nested 10-fold cross-validation for removing redundant predictors. The ROIs with large positive weights in SVM and left after lasso regularisation (with low cross-validation error) were considered as risk factors. These positive-weighted ROIs were linked with AUD individuals who relapsed within 6 months, and ROIs with large negative weights were protective factors contributing to abstinence.
Different neural representation of VOR and RA between AUD and healthy individuals
Findings
Different neural representation between AUD and healthy individuals
In VOR (RDM-fc8), motor-related brain areas (precentral and supplementary motor cortex) and postcentral cortex of AUD individuals showed higher involvement than in healthy individuals (figure 2A, table 1 and online supplemental table 3). Modelling with RDM-fc7 showed similar significant regions as RDM-fc8 (see online supplemental table 4), while RDM-fc6 did not show significant difference between two groups.

{kind=link}
{kind=link}
Increased decoding neural patterns and their connections to the whole brain. (A) The increased decoding neural pattern in visual object recognition (VOR). (B) The increased decoding neural pattern in reward appraisal (RA). (C) Positive connectivity in ROI-to-voxel performed psychophysiological interaction (PPI) analyses: the lower half-circle displays the seed regions (RA 1 to 12 for the ROIs from RA model and VOR 1 and 2 for the VOR model) and the upper half displays the significant regions in the PPI analyses. The width of links corresponds to the size of seeds and significant regions. (D) Negative connectivity in ROI-to-voxel PPI analyses. For (A) and (B), two-sample t-tests were used between alcohol use disorder (AUD, n=238) and healthy (n=229) individuals, combining voxel-wise-p<0.0005 and FWEc=108 voxels for A and 88 voxels for B, corresponding to cluster-pFWE <0.05. For (C) and (D), one-sample t-tests were used within AUD group with a same multicomparison-correction approach for cluster-pFWE <0.05.
With RA modelling, AUD individuals showed different neural patterns in a large network compared with healthy individuals. Engaged areas involved the basal ganglia (caudate, putamen), the frontal cortex (inferior, middle and superior), precentral/postcentral gyrus and also regions in occipital and temporal cortex (figure 2B, table 1 and online supplemental table 3).
Connectivity from the neural patterns of VOR and RA
The seed-to-voxel PPI analyses were based on the enhanced decoding regions of AUD individuals compared to healthy participants, which were identified with VOR (RDM-fc8) and RA models. Supplementary motor area was positively connected to precentral/postcentral and opercular cortex during visual cue recognition in AUD individuals. During appraising reward value, a large network related to habit/reward function and executive function, as well as visual–sensory–motor processes, was involved (figure 2C,D and online supplemental table 5).
Correlation to psychometrics and relapse prediction
The involvement of neural patterns in visual recognition was positively correlated to compulsive drinking, the severity of AUD and also the AUQ score (table 2). The enhanced neural pattern in RA was correlated to the scores of AUD and AUQ. With SVM, the relapse was predicted by decoding involvements in the 14 ROIs with a balanced accuracy of 0.6220 (sensitivity=0.9583, specificity=0.2857 and parameter C=1). With lasso regularisation, four predictors (RA6, RA8, RA9 and RA10) entered the logistic regression model in most of the folds, and the balanced accuracy was 0.5104 (sensitivity=0.7027 and specificity=0.3182). See online supplemental figure 6 for the receiver operating characteristic curves). In the SVM model, the most informative ROIs (whose weights were over the mean weight) were RA6, RA 10 and RA 11, mainly including frontal eye fields (FEF), dorsolateral prefrontal cortex (dlPFC), associative visual cortex and lingual gyrus (see online supplemental table 6 for weights of all ROIs).
Correlation of averaged t-value in the ROIs and the clinical variables in all participants
Discussion
Most studies of cue reactivity in the past decades have not disentangled the processing of cues into components. For the first time, the current study comparing AUD individuals and healthy participants found specific enhanced patterns in VOR and RA of alcohol cues, as well as their relevance for clinical characteristics and outcome. Furthermore, we found the neural patterns connected to large-scale functional networks, and the decoding involvements of enhanced neural patterns could contribute to predicting relapse within 6 months.
Role of sensory and motor regions
Comparing the neural representation between AUD and healthy individuals, we found the sensory and motor system of AUD individuals had enhanced information decoding in both visual VOR and RA processes. Some recent neuroimaging studies found sensory and motor function could also be relevant in the development of addiction.6 15 An animal study in 2022 reported that occipital cortical areas lost their specific interaction with sensory insular cortex, striatal and sensorimotor networks after chronic alcohol consumption, because of a regional increase in neuronal activity and overall correlation.7
The visual cortex is the first gate for visual cues in the cortex. Studies from animals and humans demonstrated that both primary and higher visual cortices exhibited value-based modulations of their activity responding to reward-mounted cues.6 In our RA model of RSA, we observed the enhanced neural pattern located at higher visual cortices (BA18, BA19 and BA21), which means that the visual cortex of AUD individuals might represent the reward value of alcohol cues better than in healthy individuals. Interestingly, it was observed in the VOR processing that the somatosensory cortex, which is mainly responsible for low-level tactile information, was also involved in the specific neural pattern in AUD individuals. A possible explanation might be that alcohol had a powerful impact on the somatosensory circuits and the exposure to visual cues may meanwhile activate sensory representations in the haptic modality. In PPI analysis, fusiform gyrus and associative visual cortex (V3, V4 and V5) were connected to striatum (putamen and caudate), and this could be related to value modulation on visual recognition (top–down influences).16
In both VOR and RA processes, AUD individuals showed special neural representation in motor and premotor brain areas. One interpretation could be that the motor brain regions play a role in the formation of automatised drinking behaviour, which is also known as habitual and compulsive drinking.4
The positive correlation to OCDS scores could also support this interpretation. Especially VOR-related patterns were associated with compulsive drinking, whereas RA was only associated with the severity of dependence and craving. Habitual drinking has been explained with a concept of incentive habits,17 which is mainly related to the dorsolateral striatum. However, our findings in motor areas (also some regions in the cerebellum) might be related to automatised action schemata,6 18 as a complement to incentive habits, which would match with previous findings that heavier substance users showed a more automatised consumption.19 In many cue reactivity studies, motor-related areas also have been reported that activated differently towards substance‐related stimuli.11 15 Here we would like to specifically note that the motor brain not only represents the reward values from visual analog scale scores but also the cue features from computer vision (dCNN model), which was not reward-mounted in our hypothesis. This might imply that abnormal neural representation might be embedded deeply in the action schemata of AUD and could be crucial for therapy. Previous studies targeting automatic action tendencies showed improvements in treatment outcomes in AUD.20
From sensory to distributed processing
In addition to enhanced neural patterns in the sensory cortex, AUD individuals also showed representation in the middle temporal gyrus. The middle temporal gyrus, as a part of the associate visual cortex, plays a role as the transit hub of visual attention pathway, whose activity might reflect the sensory analysis of the cue.21 Meanwhile, our PPI analysis found a connectivity from middle temporal gyrus to the dorsal frontoparietal network (superior parietal lobule), which is involved in top–down control of visual attention.
When modelling the representation of the reward value, a large network showed high involvement in AUD individuals compared to healthy participants, especially the orbitofrontal, anterior prefrontal, dorsolateral prefrontal cortex and striatum. These areas could be summarised as three large-scale networks: reward, habit and executive, which were reviewed by Zilverstand et al in 2018 based on the impaired response inhibition and salience attribution (iRISA) model in addiction research.8 Previous studies of substance use disorder demonstrated that the hyperactivation of these three networks was involved in the appraisal of the subjective value of the salient cues, automatisation of the reaction and cognitive control towards processing the cues. Moreover, when drug incentives elicited stronger activation in these three networks in individuals with substance use disorder compared with controls, the aberrant salience attribution to drug-related stimuli was found to interact with impaired response inhibition in drug addiction.5 8 9 22 Our recent work suggested that the interaction of these three key networks may be rebalanced by an opioid receptor antagonist.23
It is indicated that even the passive-viewing cue exposure could involve key addiction networks beyond value appraisal, habit learning and response inhibition24 to higher cognitive processes.25 Using PPI we found that AUD individuals showed increased connectivity from higher visual cortex and attention system to higher cognitive functioning-related area, including regions of dorsolateral prefrontal cortex, supramarginal, angular gyri and anterior prefrontal cortex, as well as the memory system (hippocampus and parahippocampal gyrus). The supramarginal and angular gyri were found as the crucial regions for active maintenance of information in a working memory task.26 Besides, the connectivity from visual and attention system to anterior and dorsolateral prefrontal cortex could also reflect the information transfer towards value circuit executive system, which supports goal-directed behaviour, inhibitory control and also self-regulatory processes.27
Limitations
We were not able to investigate the temporal characteristics of the neural pattern because of the design of paradigm and the temporal resolution. To answer questions such as whether the VOR pattern responds to cues before RA pattern or synchronously, a long-event fMRI paradigm design could be used in the future, combining with finite impulse response modelling or maybe magnetoencephalography/electroencephalography could also be used to characterise the temporal characters of the neural pattern. The prediction models, both SVM and lasso penalised logistic regression, did not reach a higher accuracy compared with a previous study, which predicted relapse with cue-reactivity fMRI and structural MRI.28 In this study, we focused on neural cue processing, which might be not sufficient as the only predictor. Although this study had a relatively large sample size, the sample size for participants with relapse data was small. Another limitation of using the lasso penalised logistic regression might be the relatively weak linear association between neural patterns and relapse.
Clinical implications
With linear SVM, our results showed that the decoding involvement of enhanced neural patterns of cue reactivity could contribute to predicting relapse within 6 months. Although the accuracy of prediction was moderate, it might still imply a potential neuroimaging marker for clinical practice. The high-weighted features in the SVM model, the decoding involvements of FEF and dlPFC had large negative weights towards relapse, which could be considered as protective factors. The FEF and dlPFC could be related to the attention and executive function (dorsal frontoparietal network), and by targeting the dorsal frontoparietal network, attentional bias modification therapy has been studied in substance use disorder.29 Moreover, transcranial magnetic stimulation with the target area of dlPFC was reported modulating neural activity in brain circuits that mediate cognitive processes relevant to addiction,30 and the findings in our current study could suggest further potential target regions.
Conclusions
In this study, we found enhanced neural representation of alcohol cues in specific brain regions of AUD individuals in the context of VOR and RA. Furthermore, we found small-to-moderate associations between neural patterns and clinical measures and relapse. The identification of these dysfunctional processes of cue reactivity in AUD individuals might bring a deeper understanding of the neural and psychological mechanisms underlying AUD and could be an important step towards the goal of precision medicine approaches in AUD.
Data availability statement
Data are available on reasonable request.
Ethics approval
This study involves human participants. All projects in this study were approved by the local ethics committee of the University of Heidelberg (ethics approval nos: 2007-095F-MA, 2009-215N-MA, 2010-348N-MA, 2011-303N-MA, 2009-215N-MA and 2015-540N-MA). Participants gave informed consent to participate in the study before taking part.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors HT and SV-K were responsible for the study design, and SV-K is responsible for the overall content as guarantor. HT performed the data analysis. MFG, PB, OH, PK and SV-K supervised the analysis. HT, MFG, PB, WS and SV-K contributed to interpretation of data. HT drafted the manuscript. AML revised the manuscript as a native English speaker. WS, PK, FK and SV-K procured study funding. All authors revised the manuscript critically for important intellectual content and approved the final version.
Funding This study was partly supported by grants from the Deutsche Forschungsgemeinschaft (SFB 636/D6, TRR 265 Project ID-402170461, Project ID-421888313, Project-ID 437718741 and GRK2350-1 Project-ID 324164820); the European Union’s ERA-Net NEURON and Horizon 2020 programs (FKZ 01EW1112-TRANSALC and 668863-SyBil-AA); the Federal Ministry of Education and Research (Project ID 01ZX1909 and FKZ-01GS08152) and by an investigator-initiated research grant supported by Lundbeck A/S, Denmark. We would like to thank Professor Rainer Spanagel, Dr Anita Hansson, Dr Anne Koopmann and Professor Karl Mann for the provision of fMRI and clinical data of the included previous studies, and Maria Bernardo for her advices on machine learning approaches, as well as Sabine Hoffmann for her assistance with data management.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.